DNA -replisomet: Et design-paradigme (Understreking og kursiv, ved oversetter.)
Av Jonathan McLatchie, 21. mars 2024. Oversatt herfra
Bilde 1. Helikase
DNA -replisomet er en av de mest bemerkelsesverdige molekylære maskinene, og involverer et kompleks av forskjellige proteiner, som hver er veldig spesifisert for å oppfylle sin rolle i prosessen med å replisere genomet i forberedelsen til celledeling. Hastigheten for DNA-replikasjon er blitt målt til hele 749 nukleotider per sekund(1), og feilhastigheten for nøyaktige polymeraser antas å være i området 10-7 til 10-8, basert på studier av E. coli og bakteriofag DNA-replikasjon.(2)
En av de beste animasjonene av denne utrolige prosessen er denne av den australske animatøren Drew Berry:
Det er vanskelig å se på en animasjon som denne (som er drastisk-overforenklet) og ikke komme ut med den sterke intuisjonen at en så intrikat koreografert maskin, er et produkt av mesterlig ingeniørfag. Stabile og funksjonelle proteinstrukturer er astronomisk sjeldne i kombinatorisk sekvensrom, og DNA -replikasjon krever mange av dem. Men ikke bare noen gamle stabilt sammenleggbare proteiner vil gjøre jobben. Disse proteinene må lages veldig spesielt for å utføre sine respektive jobber. Når man fokuserer på spesifikke proteiner, tar det faktisk design intuisjonen til nye høyder. Se for eksempel disse vakre animasjonene av topoisomerase -lenke, helikase lenke og DNA -polymerase lenke. Én artikkel oppsummerer den tekniske dyktigheten av DNA -replikasjon slik:
 "Syntese av alt genomisk DNA involverer den sterkt koordinerte virkningen av flere polypeptider. Disse proteinene samler to nye DNA -kjeder i et bemerkelsesverdig tempo, og nærmer seg 1000 nukleotider per sekund i E. coli. Hvis DNA -dupleksen var 1 m i diameter, vil følgende utsagn grovt beskrive E. coli -replikasjon. Gaffelen ville bevege seg på omtrent 600 km/t, og replikasjonsmaskineriet ville være omtrent på størrelse med en FedEx leveringsbil. Replikering av E. coli -genomet ville være en 400 km tur for to slike maskiner, noe som i gjennomsnitt vil gjøre en feil bare en gang hver 170 km. Den mekaniske dyktigheten til dette komplekset er enda mer imponerende gitt at det syntetiserer to kjeder samtidig når det beveger seg. Selv om den ene tråden syntetiseres i samme retning som gaffelen beveger seg, blir den andre kjeden (den hengende tråden) syntetisert på en stykkevis måte (som Okazaki -fragmenter) og i motsatt retning av total gaffelbevegelse. Som et resultat, omtrent en gang i sekundet, må en leveringsperson (dvs. polymerase aktivt sted) tilknyttet lastebilen ta en omvei, komme av, og deretter gå sammen med malen DNA -tråden for å syntetisere 0,2 km fragmenter." (3)
"Syntese av alt genomisk DNA involverer den sterkt koordinerte virkningen av flere polypeptider. Disse proteinene samler to nye DNA -kjeder i et bemerkelsesverdig tempo, og nærmer seg 1000 nukleotider per sekund i E. coli. Hvis DNA -dupleksen var 1 m i diameter, vil følgende utsagn grovt beskrive E. coli -replikasjon. Gaffelen ville bevege seg på omtrent 600 km/t, og replikasjonsmaskineriet ville være omtrent på størrelse med en FedEx leveringsbil. Replikering av E. coli -genomet ville være en 400 km tur for to slike maskiner, noe som i gjennomsnitt vil gjøre en feil bare en gang hver 170 km. Den mekaniske dyktigheten til dette komplekset er enda mer imponerende gitt at det syntetiserer to kjeder samtidig når det beveger seg. Selv om den ene tråden syntetiseres i samme retning som gaffelen beveger seg, blir den andre kjeden (den hengende tråden) syntetisert på en stykkevis måte (som Okazaki -fragmenter) og i motsatt retning av total gaffelbevegelse. Som et resultat, omtrent en gang i sekundet, må en leveringsperson (dvs. polymerase aktivt sted) tilknyttet lastebilen ta en omvei, komme av, og deretter gå sammen med malen DNA -tråden for å syntetisere 0,2 km fragmenter." (3)
Bilde 2. Oppbygning av Topoisomerase
Irreduserbar kompleksitet på steroider
DNA -replikasjon er et eksempel på hva vi kan kalle "irreduserbar kompleksitet på steroider." Genom duplisering er en forutsetning for differensiell overlevelse, som er nødvendig for prosessen med naturlig seleksjon for til og med å arbeide. Dermed kan man knapt appellere til naturlig utvalg for å redegjøre for opprinnelsen til DNA -replikasjon uten å anta eksistensen av tingen man prøver å forklare. Det er vanskelig å se for seg et levedyktig replikasjonssystem som er enklere enn DNA -replisomet vist i animasjonen ovenfor. Selv om RNA World-scenariet (som fastholder at RNA-basert liv går forut for liv basert på DNA og proteiner) er en populær hypotese, florerer problemer for dette scenariet, slik det har blitt diskutert mange ganger her i Evolution News og i forskjellige andre publikasjoner (f.eks. i Stephen Meyers bok, Signature in the Cell, kapittel 14-lenke). For eksempel er en av de fremste  utfordringene den iboende ustabiliteten til RNA (å være enkeltstrenget og ha en ekstra 2 'OH -gruppe, noe som gjør den utsatt for hydrolyse). RNA -polymerer er derfor ekstremt usannsynlig å ha overlevd i det tidlige jordmiljøet lenge nok til å ha verdi. For det andre, når RNA danner komplementære basepar for å brette tilbake på seg selv, presenterer deler av molekylet ikke lenger en eksponert streng, som kan tjene som en mal for kopiering. Dermed er det en fysisk begrensning for muligheten for RNA til selvreplikering.
utfordringene den iboende ustabiliteten til RNA (å være enkeltstrenget og ha en ekstra 2 'OH -gruppe, noe som gjør den utsatt for hydrolyse). RNA -polymerer er derfor ekstremt usannsynlig å ha overlevd i det tidlige jordmiljøet lenge nok til å ha verdi. For det andre, når RNA danner komplementære basepar for å brette tilbake på seg selv, presenterer deler av molekylet ikke lenger en eksponert streng, som kan tjene som en mal for kopiering. Dermed er det en fysisk begrensning for muligheten for RNA til selvreplikering.
En ytterligere grunn til at DNA-replikasjonsmaskineriet viser irreduserbar kompleksitet på steroider, er at det, som er så primitivt, det er langt mer vanskelig å se for seg noen form for kooperasjonsscenario, enn det ville være for et system som oppsto mye senere, for eksempel det bakterielle flagellum . Med flagellen kan man i det minste peke på alternative funksjoner som kan utføres av et antall av flagellære komponenter (for eksempel type III sekresjonssystem). Imidlertid, med DNA-replikasjon, er det uklart hvilke andre systemer noen av komponentene kan være kooperert fra, siden ethvert annet system ville ha oppstått etter opprinnelsen til DNA-replikasjon.
En enda mer slående gåte er at de viktigste enzymer (spesielt replikative polymeraser) ikke er homologe, noe som har ført til antydningen om at DNA -replikering kan ha oppstått mer enn én gang.(4,5) Denne observasjonen passer mer komfortabelt til et designparadigme enn til ett forpliktet på naturalisme.
Bilde 3. Interavhengige komponenter i cellen
Hvilke komponenter er essensielle for DNA -replikasjon?

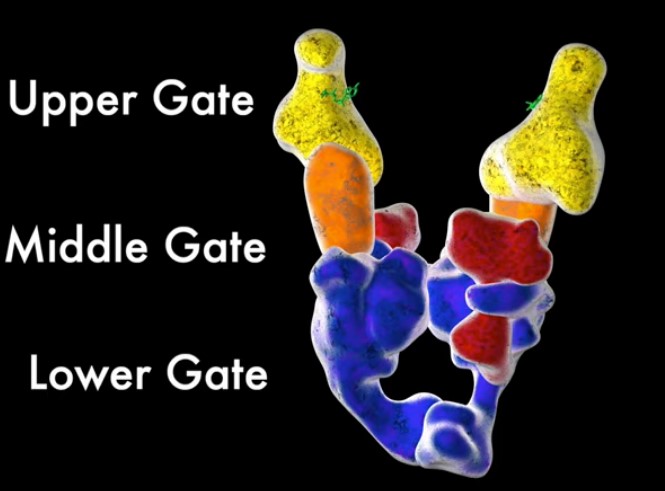
Hvilke proteinkomponenter som er involvert i DNA -replikasjon er uunnværlige for funksjon? For det første er det DNA -polymerasen som faktisk utfører kopieringen av hver strand. Uten det ville ingen replikasjon funnet sted i det hele tatt. Men DNA -polymerasen klarer ikke å begynne replikasjon uten tilstedeværelse av en fri 3 'OH (hydroxyl) gruppe. Dermed skaper et annet enzym - en form for RNA -polymerase kalt en primase - et kort RNA -fragment (kalt en primer) som DNA -polymerasen kan strekke seg fra (i motsetning til DNA -polymerase, krever ikke primasen tilstedeværelsen av en fri 3 'OH -gruppe ). I fravær av primase -enzymet ville ingen RNA -primere bli lagt ned på, verken den ledende eller hengende tråden, og DNA -replikasjon ville ikke være i stand til å kommunisere. Videre må selve DNA-polymerasen festes til DNAet med et ringformet protein, kjent som en glideklemme (som forhindrer at det faller av DNA-maltråden). Men glideklemmen kan ikke feste seg direkte til DNA på egen hånd. I stedet formidler et proteinkompleks som kalles klemmelasteren, belastningen av glideklemmen på DNA ved replikasjonsgaffelen, og losser energien fra ATP -hydrolyse for å åpne glideklemme og laste den på DNA. I fravær av glideklemme eller klembelasteren, ville DNA -polymerasen ofte falle av DNA -malen, noe som ville gjøre den ekstremt ineffektiv.
Replikasjonsprosessen kan selvfølgelig ikke begynne med mindre DNA -dobbelthelixen er 'pakket ut', og dette oppnås av enzymhelikasen, som bryter hydrogenbindinger langs DNA -molekylet, og dermed åpner opp og utsetter de to trådene for replikasjon av polymerasen. I dens fravær vil DNA -polymerasen stoppe og ikke være i stand til å skille trådene som ligger foran.
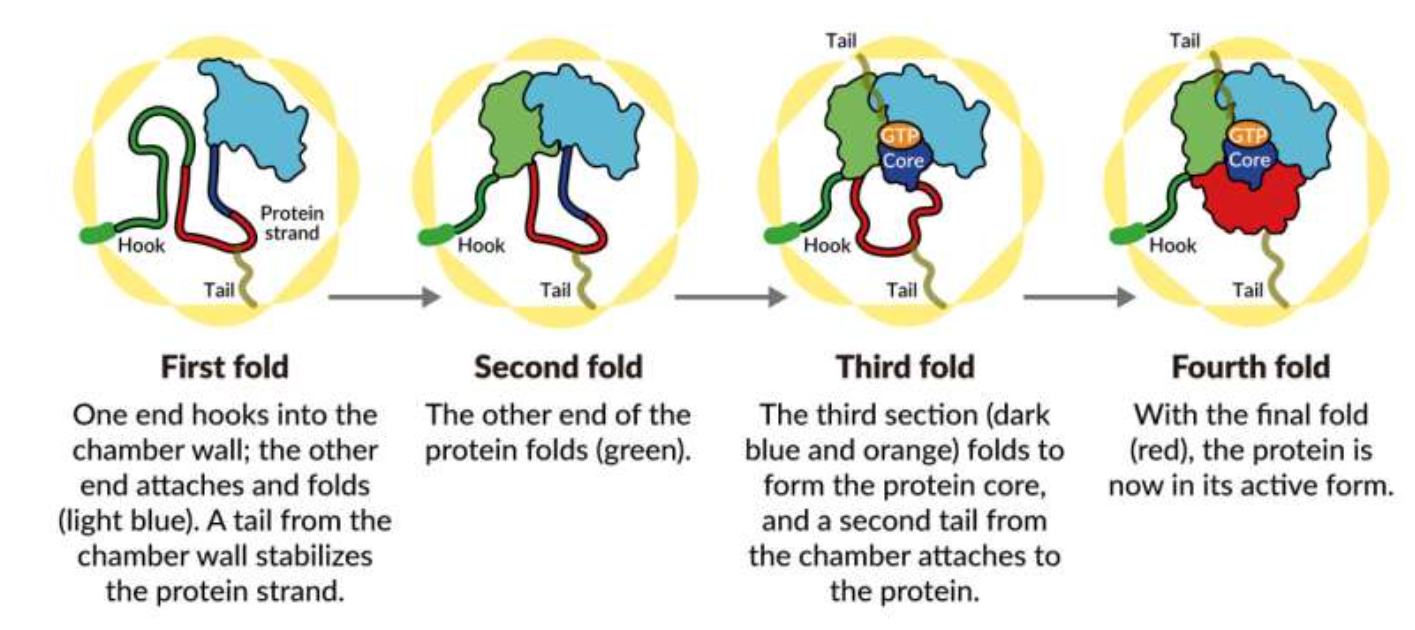
Bilde 4. Spesifisert kompleksitet -orndet i henhold til en oppskrift (mal)
Selv med helikase -enzymet som skiller de to strengene, vil tråder sannsynligvis gjenkjennesunder kopieringsprosessen. Det å gå inn i de enkeltstrengede bindende proteiner som binder seg til de utsatte DNA-båndene, forhindrer dem i å gjenkjennes nder kopiering. Uten dem ville DNA -tråden binde seg sammen igjen før de kunne kopieres.
 Topoisomerase -enzymer er nødvendige for å fjerne supercoiler som induseres av torsjonsstresset. De gjør det ved å kutte den ene tråden, passere den andre tråden gjennom gapet og deretter reforsegle bruddet. I fravær av topoisomerase -enzymer, ville DNA til slutt brytes og dermed hindre DNA -replikasjonsprosessen.
Topoisomerase -enzymer er nødvendige for å fjerne supercoiler som induseres av torsjonsstresset. De gjør det ved å kutte den ene tråden, passere den andre tråden gjennom gapet og deretter reforsegle bruddet. I fravær av topoisomerase -enzymer, ville DNA til slutt brytes og dermed hindre DNA -replikasjonsprosessen.
På grunn av den anti-parallelle naturen til DNA (og det faktum at DNA-polymerasen bare kan replikere i en 5 'til 3' retning), må en tråd, den hengende tråden, replikeres bakover (for at replikasjonsgaffelen skal bevege seg i en enkelt retning). Dette gjøres diskontinuerlig i små seksjoner. RNA -primere er lagt ned av primase, og fra disse syntetiseres korte fragmenter av DNA kjent som Okazaki -fragmenter. RNA -primerne blir deretter fjernet og erstattet med DNA, og Okazaki -fragmentene blir sydd sammen av enzymligasen. Vi har allerede diskutert nødvendigheten til primase -enzymet for å syntetisere RNA -primere. Det kan legges til at i fravær av RNA -eksisjonsenzymer (som fjerner RNA -primere), vil RNA -fragmentene forbli kovalent festet til de nylig replikerte fragmentene av DNA. I mangel av ligase (som knytter Okazaki -fragmentene sammen), vil de nylig repliserte trådene dessuten forbli som fragmenter.
Hvis fjerning av noen av de nevnte komponentene ville gjøre DNA-replikasjonsmaskineriet ikke-funksjonelt, hvordan kan et slikt system komme gjennom en trang darwinsk trinnvis vei, og bevare selektiv nytteverdi på hvert trinn underveis? Uansett hvilken prosess som ble produsert, måtte DNA -replisomet vite hvor målet var. En slik årsak må være teleologisk.
Bilde 5. Eks. på teleologisk drevet prosess
Et paradigme av design
 DNA-replikasjonsmaskineriet representerer et av de mest ekstraordinære eksemplene på nanoteknologi som finnes i cellen. I et hvilket som helst annet erfaringsområde, vil et så komplekst og delikat arrangement av deler umiddelbart bli anerkjent som å reflektere bevisst intensjon - det vil si som et produkt av et sinn. Hvorfor skal en slik slutning ikke tillates ved å undersøke biologiske systemer? For mer detaljert behandling om denne fascinerende molekylære maskinen, se intervjuet mitt om den fra sist sommer On ID Future -lenke. Jeg publiserte også en tidligere serie (for mer enn ti år siden) som utforsket de forskjellige proteinkomponentene mer detaljert. Du kan finne disse her:
DNA-replikasjonsmaskineriet representerer et av de mest ekstraordinære eksemplene på nanoteknologi som finnes i cellen. I et hvilket som helst annet erfaringsområde, vil et så komplekst og delikat arrangement av deler umiddelbart bli anerkjent som å reflektere bevisst intensjon - det vil si som et produkt av et sinn. Hvorfor skal en slik slutning ikke tillates ved å undersøke biologiske systemer? For mer detaljert behandling om denne fascinerende molekylære maskinen, se intervjuet mitt om den fra sist sommer On ID Future -lenke. Jeg publiserte også en tidligere serie (for mer enn ti år siden) som utforsket de forskjellige proteinkomponentene mer detaljert. Du kan finne disse her:
"DNA -replikering: et ingeniørmessig under":
"Replikering av DNA med ekstraordinær nøyaktighet: møt DNA -polymerasen" -lenke.
"Tvinn opp den doble helixen: møt DNA -helikasen" -lenke.
Hvis du likte animasjonen av Drew Berry i begynnelsen av denne artikkelen, er her en mer detaljert animasjon, produsert av Oxford University Press -lenke. Her er en annen animasjon som avslører hvordan DNA -polymerasene er koblet -lenke, slik at de kan bevege seg i samme retning.
Bilde 6. Trosgrunnlag for ateister
For referanser, se slutten av artikkelen her.
 JONATHAN MCLATCHIE Bilde 7.
JONATHAN MCLATCHIE Bilde 7.
RESIDENT BIOLOG & stipendiat, SENTER FOR VITENSKAP OG KULTUR
Dr. Jonathan McLatchie har en bachelorgrad i rettsmedisinsk biologi fra University of Strathclyde, en mastergrad (M.Res) i evolusjonsbiologi fra University of Glasgow, en annen mastergrad i medisinsk og molekylær biovitenskap fra Newcastle University, og en doktorgrad i evolusjonær Biologi fra Newcastle University. Tidligere var Jonathan assisterende professor i biologi ved Sattler College i Boston, Massachusetts. Jonathan har blitt intervjuet på podcaster og radioprogrammer, inkludert "Unbelievable?" på Premier Christian Radio og mange andre. Jonathan har talt internasjonalt i Europa, Nord-Amerika, Sør-Afrika og Asia for å fremme bevis på design i naturen.
Musikk til arbeidet.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund